Гидролиз пирофосфата, образующегося, в частности, как побочный продукт в результате биосинтеза таких важнейших клеточных полимеров как ДНК и РНК, до двух молекул неорганического фосфата – одна из необходимых реакций, протекающих в живых клетках как про- так и эукариотических организмов и катализируемых пирофосфатазами (PPase). Этот гидролиз сопровождается выделением значительных количеств свободной энергии, но поскольку эта реакция в большинстве случаев катализируется локализованными в цитопламе ферментами, то выделяемая энергия непродуктивно превращается в тепло и не подвергается запасанию для использования в других метаболических превращениях. Однако некоторые организмы, к которым, в частности, относятся и бактерии Rhodospirillum rubrum,наряду с типичной цитоплазматической пирофосфатазой имеют также дополнительный мембрано-связанный фермент, способный гидролизовать пирофосфат в реакции, сопряженной с экспортом протонов через бактериальную мембрану с формированием протонного потенциала, способного, как хорошо известно, в дальнейшем генерировать образование АТФ за счет «протон-движущей силы». Можно считать, что при гидролизе пирофосфата такими ферментами, например (H+-PPaseRru), выделяемая энергия не полностью расходуется в виде тепла, а частично запасается для возможных дальнейших метаболических превращений в виде протонного мембранного потенциала клетки, способного генерировать АТФ. Тем самым, можно было надеяться, что в случае генно-инженерного создания на базе таких клеток – продуцентов практически полезных метаболитов и макромолекул, требующих для своего биосинтеза больших количеств клеточной энергии, использование мембано-связанных и протон-экспортирующих пирофосфатаз окажется достаточно выгодным, поскольку, сохраняя все традиционные пути генерации энергии клеток дикого типа, такие штаммы будут иметь дополнительные возможности, обусловленные специфичностью гидролиза пирофосфата этими клетками. Такая принципиальная возможность впервые продемонстрирована в данной работе в результате молекулярного клонирования, интеграции и обеспечения экспрессии в хромосоме клеток Escherichia coli структурной части гена (H+-PPaseRru) с «гармонизированными» для трансляции в E. coli аминокислотными кодонами.
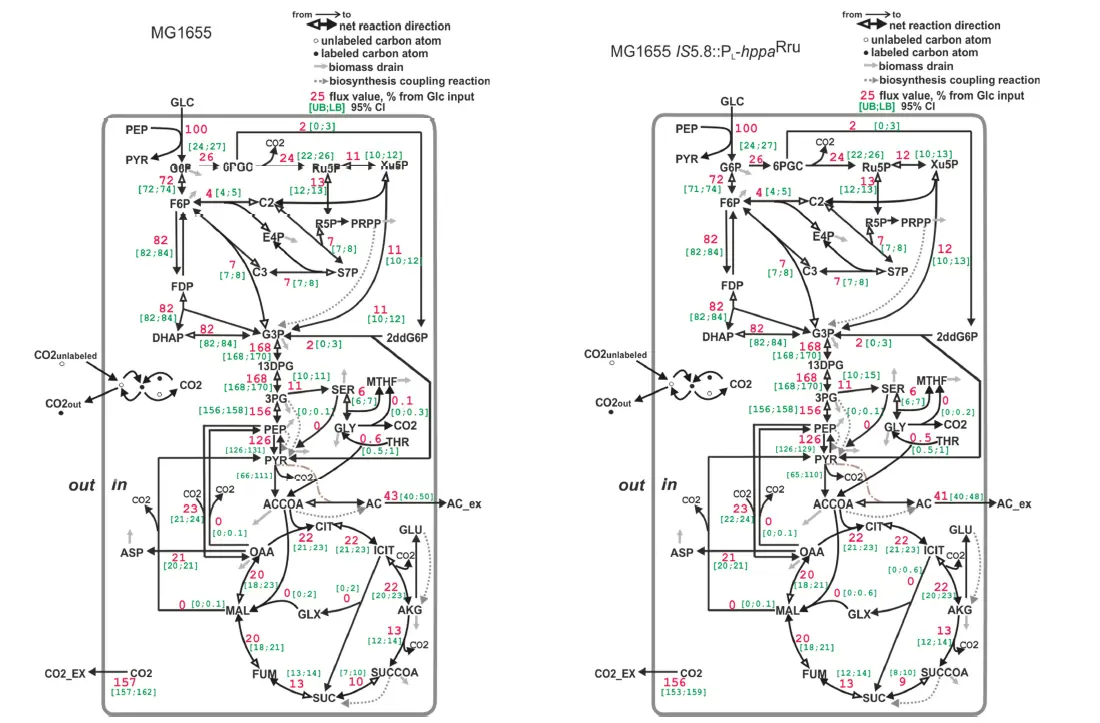
Первоначально было показано, что на фоне функционирования гетерологичного гена собственный ген цитопламатической пирофосфатазы, ранее относящийся к незаменимой для E. coli группе жизненно важных генов, может быть успешно делетирован стандартными процедурами Рекомбиниринга, хотя и со значительным замедлением роста полученных рекомбинантных E. coli штаммов. Прямое изменение распределения метаболических потоков в рекомбинантном штамме с замененным природным геном пирофосфатазы на гетерологичный было убедительно продемонстрировано прецизионным методом 13С-MFA (13C-Metabolic Flux Analysis). Действительно, было достоверно продемонстрировано (смотри Рисунок) значительное (36%) уменьшение метаболических превращений в цикле трикарбоновых кислот (ТСА) рекомбинантного штамма, что, по-видимому, связано с отсутствием потребности в генерации большого количества прямых предшественников синтеза АТР (а именно, NADH) в ТСА из-за наличия возможности синтеза АТР другим способом в рекомбинантном штамме. Таким образом перспективность использования предложенного подхода для конструирования продуцентов энергозависимых соединений впервые экспериментально продемонстрирована.
